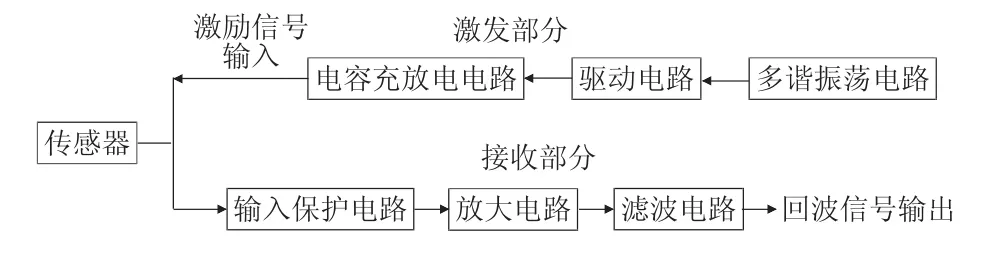
![图片[1]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172016-68753c50b903f.webp)
![图片[2]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172018-68753c52d35dc.webp)
![图片[3]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172020-68753c545cbf9.webp)
![图片[4]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172021-68753c55db27b.webp)
![图片[5]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172023-68753c5788ca8.webp)
![图片[6]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172025-68753c592cff5.webp)
![图片[7]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172026-68753c5abf141.webp)
![图片[8]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172028-68753c5c78d36.webp)
![图片[9]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172030-68753c5e270e9.webp)
![图片[10]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172031-68753c5fd8808.webp)
![图片[11]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172033-68753c61bb4e1.webp)
![图片[12]-蒺藜苜蓿MtDWF1基因克隆、亚细胞定位及表达特征分析-游戏花园](http://www.hunggame.com/wp-content/uploads/2025/07/20250714172036-68753c6426869.webp)
摘要:Delta24-甾醇还原酶(Delta24-sterol reductase,DWF1)是油菜素内酯(Brassinosteroide,BR)合成途径中的关键酶,在植物生长发育中起着关键性的作用。为了研究DWF1基因在蒺藜苜蓿(Medicago truncatula)中的功能,本试验从蒺藜苜蓿R108中克隆MtDWF1基因全长序列,该基因开放阅读框(ORF)1 704 bp,编码567个氨基酸,分子量65.911 kD,理论等电点为8.53。亚细胞定位显示,该蛋白定位于细胞质内。进化分析表明,蒺藜苜蓿MtDWF1蛋白与鹰嘴豆(Cicer arietinum)、红三叶(Trifolium pratense)及豌豆(Pisum sativum)亲缘关系较为接近。MtDWF1基因在蒺藜苜蓿根、茎、叶中均有表达,在茎和叶中的表达水平较高。外源激素脱落酸(Abscisic acid,ABA)能够明显提高MtDWF1表达,水杨酸(Salicylic acid,SA)能够显著降低MtDWF1表达水平。同时,经油菜素内酯诱导后,MtDWF1表达水平整体呈现增加趋势。本研究为进一步探索MtDWF1基因在蒺藜苜蓿发育过程中的作用提供理论研究基础。
关键词:MtDWF1;蒺藜苜蓿;基因克隆;表达分析;亚细胞定位
中图分类号:Q785文献标识码:A文章编号:1007-0435(2023)04-0984-08
Cloning,Subcellular Localization and Expression Pattern of MtDWF1 Gene in
Medicago truncatula
XIE Hong1, LI Shu-wen1, DONG Di1, WANG Meng-di JIA Wei CHAO Yue-hui ZHAO Yan
(1. School of Grassland Science, Beijing Forestry University, Beijing 100083, China; 2. College of Grassland, Resources
Environment, Inner Mongolia Agricultural University, Hottot, Inner Mongolia 010018, China)
Abstract:Delta24-sterol reductase (DWF1) is a crucial enzyme in the biosynthesis of brassinosteroids and has a significant regulation on plant growth and development. To examine the function of the DWF1 gene in Medicago truncatula,the full-length sequence of the MtDWF1 gene was cloned from M. truncatula (R108). The open reading frame of MtDWF1 was found to be 1 704 bp in length and encoded a peptide of 567 amino acids with a molecular weight of 65.911 kDa and a predicted isoelectric point of 8.53. The protein of the MtDWF1 was localized in the cytoplasm as determined by subcellular localization analysis. Evolutionary analysis indicated that the MtDWF1 protein of M. truncatula is closely related to those of Cicer arietinum,Trifolium pratense,and Pisum sativum. The MtDWF1 gene was expressed in the roots,stems,and leaves of M. truncatula,with higher expression levels observed in stems and leaves than that in roots. Exogenous application of abscisic acid (ABA) caused the MtDWF1 expression significantly increased,while salicylic acid (SA) significantly reduced. Brassinosterol (BR) treatment resulted in an overall increase of MtDWF1 expression. This study provides a foundation for further investigating on the role of the MtDWF1 gene in M. truncatula development.
Key words:MtDWF1;Medicago truncatula;Gene clone;Expression analysis;Subcellular localization
蒺藜苜蓿(Medicago truncatula),豆科(Leguminosae),苜蓿属(Medicago L.),一年生牧草植物。因其倍性小(2n=16)、基因组小(5×108bp)[1]、生长期短、自花授粉、结实率高及可固氮等优良性状,受到了越来越多研究者们的青睐[2-4],同时也成为了研究豆科生物和基因组学方面的模式植物[5]。
油菜素内酯(Brassinosteroide,BR),是一类调控植物生长的固醇类激素[6],被称为第六类植物激素,是植物生长发育所需的天然植物生长调节剂[7],在植物的不同器官中均存在,参与种子萌发[8]、细胞伸长和分裂、叶片衰老以及增加抗旱、抗寒、抗病等不同逆境环境的抵抗[9]。研究发现,DWF基因家族与BR在植物生长发育过程中联系密切,很多成员都参与了BR的合成途径[10]。在拟南芥(Arabidopsis thaliana)[11] 中DWF1/DIM是一个双功能蛋白,催化24-亚甲基胆甾醇合成油菜素内酯,在dwf1/dim突变体中,内源BR的合成受阻其含量大量降低,造成DWF1基因的表达水平降低,从而导致植株生长速度减缓,植株表现矮化现象。在种子萌发时,DWF4与BR合成相关基因的表达量上调,导致内源BR含量增加。在DWF1蛋白序列中存在FAD结合结构域,具有FAD氧化还原酶特性。此外,DWF1在植物中的其他功能也得到了充分的研究,对甜瓜(Cucumis melo)中的DWF1基因进行克隆及表达分析,结果发现能促进根系及叶等组织的发育[12]。在水稻(Oryza satival)中,dwf1突变体的植株矮小,生长速度减缓,根、茎、叶等器官明显变脆,同时叶片数量也较少,结实率及千粒重均比野生型有一定的降低[13]。在玉米(Zea mays)中通过RNAi技术干扰DWF1,发现基因表达受到抑制,转基因植株变矮,同时这种抑制与植株的高度呈现正相关[14]。
综上,DWF1基因在植株生长发育过程中发挥着重要的作用,然而在豆科模式植物蒺藜苜蓿中关于DWF1基因相关的研究较少。因此,本试验以蒺藜苜蓿为研究材料,从蒺藜苜蓿中通过克隆得到MtDWF1基因,对其进行生物信息学分析、亚细胞定位及结合qRT-PCR分析该基因的表达特征,初步探讨MtDWF1的功能,为后续深入研究该基因的功能机制提供理论基础。
1材料与方法
1.1试验材料
试验所用的野生型蒺藜苜蓿(生态型R108)种子于北京林业大学保存。蒺藜苜蓿种子经4℃春化3 d后放置于光照16 h /25℃黑暗8 h/23℃至根系发育。然后移栽至直径12 cm花盆中,并于25℃光照16 h/24℃黑暗8 h条件下生长。反转录试剂盒、PrimeSTAR MAX、限制性内切酶、pMD19-T克隆载体均来自于TAKARA公司;纯化试剂盒、DNA提取试剂盒购自于OMEGA公司;DL5000 DNA Marker购自艾克瑞公司;荧光定量TB Green订购于康维公司;大肠杆菌感受态DH5α来自于天根公司;利福平、卡那霉素、ABA,SA和BR购于Sigma公司,农杆菌感受态EHA105为实验室保存。其他试剂均采自于进口分装或国产分析纯。
1.2试验方法
1.2.1引物设计基于已经公布的蒺藜苜蓿R108参考基因组(R108_HM340_v1.0),通过软件Primer Premier 5.0进行引物设计。其中,MtDWF1-F/R用于MtDWF1基因全长的克隆,引物3302Y-MtDWF1-F/R用于构建亚细胞定位植物表达载体构建,引物3302Y-F和3302Y-R用于植物表达载体检测,引物MtDWF1-RT-F/R用于MtDWF1基因荧光定量检测,引物MtActin-F和MtActin-R用于内参基因Actin荧光定量检测。
1.2.2MtDWF1基因克隆取长势一致的蒺藜苜蓿健康叶片作为植物材料,将其放在盛有液氮的研钵中充分研磨,按照RNA提取试剂盒提取蒺藜苜蓿总RNA。以提取的总RNA为模板,利用反转录试剂盒进行反转录获得蒺藜苜蓿cDNA。以蒺藜苜蓿cDNA为模板,以DWF1-F/R为引物获得MtDWF1基因的目的条带。对PCR产物进行1%琼脂糖凝胶电泳检测,条带大小正确后,利用OMEGA纯化试剂盒进行纯化操作。纯化后的PCR产物与克隆载体pMD19-T进行连接。然后将连接产物转化到大肠杆菌感受态DH5α中,将其放在涂有氨苄抗生素的固体LB培养基上,挑取单菌落,使用引物M13F-47/R-48进行菌体PCR鉴定,用1%的琼脂糖凝胶电泳检测,选取扩增目的条带大小的PCR产物送公司进行测序。
1.2.3亚细胞载体构建以含有MtDWF1克隆质粒为模板,用3302Y-DWF1-F/R为引物进行PCR扩增。在50℃下使用无缝连接酶将纯化后的PCR产物与酶切质粒产物进行连接后转入大肠杆菌感受态,并将其菌液涂在含有50 mg·L-1卡那霉素的抗生素固体LB培养基上进行挑选,将其置于37℃恒温箱中培养8 h,挑取生长于抗生素培养基上的单菌落进行PCR扩增并取4 μL用1%的琼脂糖凝胶电泳进行检测,将条带大小符合的PCR产物送公司测序。
1.2.4农杆菌转化用CaCl2冻融法将3302Y-DWF1重构质粒转入到EHA105农杆菌的感受态中,并将其放置冰上30 min,液氮速冻1 min,37℃水浴5 min,冰上5 min,向管中加入1 mL的LB溶液,放置在28℃培养箱中振荡4 h,5 000 r·min-1,弃上清,将剩余菌液涂于含有利福平和卡那霉素抗生素的LB培养基上,28℃暗处理2 d后,挑取单菌落,利用引物3302Y-F和3302Y-R对菌落进行PCR检测,条带大小符合的产物送至公司测序,测序后将比对正确的农杆菌菌液加甘油保存于-80℃的冰箱中。
1.2.5亚细胞定位将构建好的35S::MtDWF1:GFP农杆菌菌液加到含50 mg·L-1的利福平和50 mg·L-1的卡那霉素抗生素的LB溶液中,黑暗条件下在培养箱振荡1~2 d后,至OD600为0.6~0.8;以健康生长一个月左右的烟草为材料,用注射器将菌液注射到烟草叶片下表皮细胞中,注射后将烟草放置于人工气候箱黑暗处理48 h,制作叶片玻片,将玻片倒置在共聚焦显微镜下,观察烟草叶片中GFP荧光信号的分布情况,最终确定目的基因所在位置。
1.2.6生物信息学分析从NCBI(https://www.ncbi.nlm.nih.gov/)数据库中下载MtDWF1的基因序列,将其在(https://iris.angers.inra.fr/obh/)中与蒺藜苜蓿R108基因组序列比对,获取MtDWF1的基因序列。同时于NCBI网站上对MtDWF1蛋白质进行保守结构域分析。利用DNA MAN分析MtDWF1的开放阅读框及编码氨基酸序列,用ProtPara(https://web.expasy.org/protparam/)分析蛋白质等电点和分子量,用(https://services.healthtech.dtu.dk/services.php?signalp-5.0)进行蛋白质信号肽的预测。利用SOPMA(https://prabi.ibcp.fr/htm/site/web/home)对MtDWF1蛋白一级、二级、三级结构进行预测,Cell-PLoc网站(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc/)去预测MtDWF1蛋白在细胞内的定位情况。通过MEGA 11软件构建同源蛋白系统进化树。
1.2.7MtDWF1实时荧光定量表达分析选取长势一致健康的蒺藜苜蓿植株根、茎、叶三个组织,液氮速冻后,迅速放入-80℃的冰箱,对其进行组织差异性表达分析。以李殷睿智[15]和舒思晨[16]的实验方法为参考,确定ABA,SA及BR浓度分别为10 μmol·L-1,0.5 mmol·L-1及0.1 μmol·L-1。选取长势一致的蒺藜苜蓿植株叶片,对叶表面均匀喷施1次进行激素诱导,分别于喷施激素的第0,0.5,1,3,6,9,12 h这7个不同时间点采取植株叶片组织后,立即用液氮进行速冻。随后将上述样品研磨后,分别提取植物的总RNA,并进行反转录。使用引物MtActin-F和MtActin-R作为荧光定量的内参基因,引物MtDWF1-RT-F/R按照试剂说明书进行qRT-PCR分析。反应结束后,根据2—△△Ct法[17]使用Excel 2010进行数据分析。
1.2.8数据分析通过Excel 2010对数据进行整理及图标制作,利用软件SPSS 26.0进行单因素方差分析。
2结果与分析
2.1蒺藜苜蓿MtDWF1基因的克隆
以DWF1-F/R为引物对蒺藜苜蓿cDNA进行PCR扩增。结果显示,如图1获得条带清晰、条带大小约1 700 bp,将其产物送至生物公司测序,测序结果表明所克隆的碱基序列与蒺藜苜蓿MtDWF1基因序列一致,初步表明蒺藜苜蓿MtDWF1基因克隆成功,将正确的克隆载体命名为pMD-MtDWF1。可用于后续试验。
2.2蒺藜苜蓿MtDWF1亚细胞定位分析
通过Cell-PLoc网站分析,推测MtDWF1蛋白定位于细胞质。将成功构建的35S::MtDWF1:GFP载体转化烟草,黑暗培养48 h,使用激光聚焦显微镜对转化后的烟草进行观察,捕捉激光信号。结果显示从细胞质捕获到强烈的GFP信号(图2)。表明MtDWF1蛋白的亚细胞定位在细胞质中。
2.3MtDWF1基因的生物信息学分析
通过PCR克隆技术得到的MtDWF1基因开放阅读框1 704 bp,编码567个氨基酸(图3),利用NCBI网站以及Blast工具对MtDWF1进行同源比对,对MtDWF1蛋白的保守结构及特性进行分析预测,发现在109~199位氨基酸之间含有FAD-binding-4结构域(图4),该位点是FAD结合位点,MtDWF1其分子式为C3018H4620N788O829S22,分子总质量65.911 kD,理论等电点为8.53。其正电荷残基数量为78,负电荷残基数量为73;不稳定系数是36.23,经预测,其蛋白性质稳定。二级及三级结构预测(图5),结果显示,MtDWF1蛋白主要由α螺旋及无规则卷曲构成,其分别各占二级结构总比例的42.5%和36.33%,含有延伸链16.23%和β转角4.94%。通过Signal 5.0网站,进行信号肽预测(图6),结果显示,MtDWF1蛋白中不含有信号肽。
亚细胞定位预测结果显示,MtDWF1蛋白定位于细胞质。使用MEGA 11软件对MtDWF1蛋白构建系统发育进化树,结果表明(图7)蒺藜苜蓿MtDWF1蛋白与鹰嘴豆(Cicer arietinum)、红三叶(Trifolium pratense)和豌豆(Pisum sativum)亲缘关系较近,与蒺藜苜蓿MtDWF1蛋白同源的物种大多为豆科植物。
2.4组织差异性分析
对蒺藜苜蓿根茎叶进行荧光定量分析,检测MtDWF1基因在根茎叶中的表达水平。结果表明MtDWF1基因在根茎叶中均有表达,在茎和叶中的表达量显著高于根,分别是根的1.45和1.46倍。MtDWF1基因在茎和叶的表达水平相似(图8)。
为探究激素对MtDWF1基因的作用,我们检测了MtDWF1在ABA,SA及BR处理下0,0.5,1,3,6,9,12 h的表达水平。研究表明,ABA处理后,MtDWF1表达量与对照相比呈现先增加后降低的趋势。0.5 h时表达量呈现逐渐增加趋势,并在6 h达到顶峰,此时MtDWF1的表达量是对照的4.81倍。随后MtDWF1表达量逐渐降低,但总体表达量高于对照(图9)。这些数据表明,ABA可促进MtDWF1的表达,促进作用在短期内较明显。
SA诱导后,MtDWF1的表达量整体呈现下降趋势,前6 h呈现急速下降趋势,在6 h时达到最低,占对照组的4.5%,6 h之后,表达量有所回升,但是仍然低于对照组。12 h表达量占对照组的44.7%(图10)。结果表明,MtDWF1对SA较敏感,且SA对MtDWF1的表达具有明显的抑制作用。
BR诱导后,MtDWF1被快速诱导其表达量整体呈现上升趋势。在处理9 h时表达水平达到峰值,其表达量是对照组的5倍(图11)。结果表明,BR对MtDWF1的表达具有促进作用。
3讨论
DWF1基因是BR合成过程中的关键基因,能够多次参与BR的催化合成。目前,DWF1基因已在多种植物中被研究,但在蒺藜苜蓿中DWF1方面的研究还未见到报道。本研究成功在蒺藜苜蓿中克隆获得MtDWF1基因,该基因全长1 704 bp,共编码567个氨基酸。进化树结果分析(图8)该基因控制合成的蛋白质与鹰嘴豆、豌豆、红三叶等多个物种中的DWF1蛋白高度同源,其与鹰嘴豆相似度最高。DWF1蛋白含有FAD-binding-4结构域保守区域。在已报道的拟南芥[18]、甜瓜(Cucumis melo)[12]、棉花(Gossypium hirsutum)[19]、马铃薯(Solanum tuberosum)[20]等物种中,DWF1主要都是参与BR的催化合成过程,同时在本试验中我们预测的蒺藜苜蓿DWF1蛋白质性质与以上这些物种中的DWF1蛋白质性质相同。因此,我们推测MtDWF1在蒺藜苜蓿中也具有相似的功能,能够参与蒺藜苜蓿BR的催化合成过程。
通过表达分析,发现MtDWF1在根、茎、叶组织中均有表达,说明其广泛存在于蒺藜苜蓿各个组织中,但其在不同的组织中的表达量具有差异性,在根中表达量最低,在叶中的表达量最高。这与DWF1基因在马铃薯[20]、玉米[14]和豌豆[21]的表达模式十分相似,推测DWF1基因在不同物种中可能具有相似的功能,因此可以利用在其他植物中获得的结果,推测MtDWF1基因可能参与了蒺藜苜蓿BR的催化合成过程。植物激素在植物生长发育过程中扮演着重要的角色,不同激素处理对基因的响应也存在着差异。ABA被称为“逆境激素”[22],在逆境胁迫下大量的脱落酸(Abscisic acid,ABA)在植物中被积累。已有研究发现,玉米在外源喷施ABA激素后DWF4的相对表达量升高[23]。在本研究中,MtDWF1受ABA激素诱导后表达量也上调,因此我们猜测,该基因表达在受到ABA诱导后上调,MtDWF1可能会对逆境胁迫产生响应,这种响应可能是受到或部分受到ABA途径所影响。除了ABA,MtDWF1与其他激素也存在相互作用。在SA处理后,该基因的相对表达量随处理时间的增加基因的表达量受到抑制,暗示SA可能对MtDWF1进行负调控从而实现对蒺藜苜蓿生长的调控。Vriet等[24]发现,DWF1基因能够促进油菜素内酯的合成,同时Best等人[25]也发现DWF1基因参与着BR和GA信号的相互作用,从而调控植物发育。在本试验中我们通过BR激素处理后,发现MtDWF1基因的表达量总体呈现上升趋势,这说明在外施BR激素的条件下,其促进了MtDWF1基因的表达,因此我们有理由猜测MtDWF1基因可能参与了蒺藜苜蓿植株的生长发育过程[20],对植株生长具有促进作用。综上所述,本研究为进一步探究蒺藜苜蓿MtDWF1基因功能提供了帮助,但关于其在蒺藜苜蓿中的具体功能及调控机制还需深入研究。
4结论
本研究成功克隆了蒺藜苜蓿MtDWF1基因并对其进行生物信息学分析显示,DWF1的开放阅读框1 704 bp,编码567个氨基酸。DWF1的序列上有一个典型的氧化还原酶的FAD保守结构域,无信号肽。亚细胞定位显示该蛋白定位于细胞质。表达分析结果表明,蒺藜苜蓿MtDWF1基因在茎及叶中的表达量最高,说明其在茎和叶中发挥作用。其表达受ABA,SA调节。BR可促进MtDWF1基因的表达,进而促进植株发育。总之,这些结果不仅为进一步探索MtDWF1基因功能的提供了思路,也从分子水平上研究其他豆科植物基因功能提供理论基础。
参考文献
[1]TRIEU A T,HARRISON M J. Rapid transformation of Medicago truncatula:regeneration via shoot organogenesis[J]. Plant Cell Reports,1996,16(1-2):6-11
[2]VIKER K B,STEELE M B,LANKOV I D,et al. Preclinical safety assessment of MV-s-NAP,a novel oncolytic measles virus strain armed with an H. pylori immunostimulatory bacterial transgene[J]. Molecular Therapy – Methods amp; Clinical Development,2022(26):532-546
[3]ENDRE G,KERESZT A,KEVEI Z,et al. A receptor kinase gene regulating symbiotic nodule development[J]. Nature,2002(417):962-966
[4]李菲,何小红,张习敏,等. 苜蓿14-3-3基因家族的鉴定与进化和特征分析[J]. 基因组学与应用生物学,2017,36(12):5238-5243
[5]TAO Y Z,ZHENG J,XU Z M,et al. Functional analysis of ZmDWF1,a maize homolog of the Arabidopsis brassinosteroids biosynthetic DWF1/DIM gene[J]. Plant Science,2004,167(4):743-751
[6]何宇炯,徐如涓,赵毓橘,等. 表油菜素内酯对油菜幼苗生长及其可溶性糖和蛋白质含量的影响[J]. 植物生理学通讯,1995,31(1):37-39
[7]曲善民,苏一诺,苗阳阳,等. 芸苔素内酯对连作及非连作紫花苜蓿生产调控初探[J]. 草地学报,2019,27(4):874-881
[8]寇江涛. 外源2,4-表油菜素内酯对NaCl胁迫下燕麦种子萌发和生理的影响[J]. 草地学报,2019,27(6):1562-1568
[9]邓天福,吴艳兵,李广领,等. 油菜素内酯提高植物抗逆性研究进展[J]. 广东农业科学,2009(11):21-25
[10]WANG Z Y,BAI M Y,OH E,et al. Brassinosteroid signaling network and regulation of photomorphogenesis[J]. Annual Review of Genetics,2012,46(1):701-724
[11]HOSSAIN Z,MCGARVEY B,AMYOT L,et al. Diminuto 1 affects the lignin profile and secondary cell wall formation in Arabidopsis[J]. Planta,2012,235(3):485-498
[12]张瑾,金乌云,陈莹莹,等. 甜瓜CmDWF1基因的克隆及表达分析[J]. 华北农学报,2019,34(5):8-14
[13]吴国超. 水稻矮脆突变体dwf1的特性与基因定位[D]. 重庆:西南大学,2014:35-36
[14]陶亚忠. 玉米与油菜素类固醇生物合成相关的基因ZmDWF1的克隆及其功能分析[D]. 北京:中国农业大学,2004:108
[15]LI Y R Z,WANG M D,GUO T,et al. Overexpression of abscisic acid-insensitive gene ABI4 from Medicago truncatula,which could interact with ABA2,improved plant cold tolerance mediated by ABA signaling[J]. Frontiers in Plant Science,2022,13(13):982715
[16]舒思晨,王娟,王柏柯,等. 外源MeJA和BR对番茄种子萌发及幼根生长的影响[J]. 农业工程,2022,12(3):138-143
[17]吴婧,范菠菠,张学峰,等. 蒙古冰草ERF转录因子生物信息学及其表达分析[J]. 草地学报,2022,30(11):2910-2921
[18]CHOE S,DILKES B P,GREGORY B D,et al. The Arabidopsis dwarf1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis[J]. Plant Physiology,1999,119(3):897-908
[19]罗明,周建平,肖月华,等. 棉花油菜素类固醇合成酶基因GhDWF1的克隆和表达分析[J]. 中国农业科学,2007(7):1337-1344
[20]唐晓,邓孟胜,邹雪,等. 马铃薯StDWF1基因克隆及表达分析[J]. 浙江农业学报,2018,30(6):909-917
[21]SCHULTZ L,HUUB L,KERCKHOFFS J,et al. Molecular characterization of the brassinosteroid-deficient Ikb mutant in pea[J]. Plant Molecular Biology,2001,47(4):491-508
[22]刚爽. ABA对亚高温强光胁迫下番茄叶片光合作用影响及分子机制研究[D]. 沈阳:沈阳农业大学,2016:9-11
[23]段方猛,罗秋兰,鲁雪莉,等. 玉米油菜素甾醇生物合成关键酶基因ZmCYP90B1的克隆及其对逆境胁迫的响应[J]. 作物学报,2018,44(3):343-356
[24]VRIET C,RUSSINOVA E,REUZEAU C,et al. From squalene to brassinolide:the steroid metabolic and signaling pathways across the plant kingdom[J]. Molecular Plant,2013,6(6):1738-1757
[25]BEST N B,HARTWIG T,BUDKA J,et al. Nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis gene DWARF1,identifying developmental interactions between brassinosteroids and gibberellins[J]. Plant Physiology,2016,171(4):2633-2647
(责任编辑 刘婷婷)